Механизмы исправления ошибок во время репликации ДНК и ее репарация вследствие повреждений на протяжении всего жизненного цикла клетки.
Основные моменты:
-
Клетки имеют различные механизмы предотвращения возникновения мутаций – необратимых изменений в ДНК
-
В процессе синтеза ДНК, большинство ДНК-полимераз «проверяют свою работу» и проводят замену бо́льшей части ошибочно вставленных нуклеотидов. Этот процесс можно назвать исправлением ошибок.
-
Сразу после синтеза ДНК любые оставшиеся ошибочные нуклеотиды обнаруживаются и заменяются в так называемом процессе репарации ошибочно спаренных нуклеотидов.
-
Если ДНК повреждена, она может быть восстановлена с помощью различных механизмов, например, путём прямой репарации, эксцизионной репарации или путём восстановления двухцепочечных разрывов
- пострепликативной репарации.
Введение
Как ДНК связана с раком? Рак возникает при неконтролируемом делении клеток, когда игнорируются клеточные «стоп»-сигналы, что приводит к образованию опухоли. Это неправильное поведение клеток вызвано накопившимися мутациями — необратимыми изменениями последовательности ДНК клетки.
На самом деле, ошибки в процессе репликации и повреждения ДНК возникают в клетках нашего тела постоянно. Однако в большинстве случаев они не приводят к раку и даже не вызывают мутаций, такие ошибки обычно обнаруживаются и исправляются в процессе репарации ДНК. Если же повреждение исправить не удаётся, то в клетке включается механизм самоуничтожения — (апоптоз), который предотвращает передачу поврежденной ДНК дочерним клеткам.
Мутации возникают и передаются дочерним клеткам только тогда, когда эти механизмы не справляются. В частности, рак возникает в случае накопившихся в одной клетке мутаций генов, связанных с делением.
В этой статье мы подробно рассмотрим механизмы, используемые клетками для исправления ошибок, которые возникают в процессе репликации. К ним относятся:
-
Исправление ошибок – процесс, который возникает во время репликации ДНК.
-
Репарация ошибочно спаренных нуклеотидов, которая происходит сразу же после репликации ДНК.
-
Механизмы репарации, которые выявляют и исправляют повреждения ДНК на протяжении всего клеточного цикла
Исправление ошибок
ДНК-полимеразы — это ферменты, участвующие в репликации ДНК. Во время копирования ДНК большинство ДНК-полимераз «проверяют», корректный ли нуклеотид они добавляют. Этот процесс называется исправлением ошибок. Если полимераза обнаружит, что был добавлен неправильный нуклеотид, она сразу же удалит и заменит его и только после этого продолжит синтез ДНКstart superscript, 1, end superscript.
Репарация ошибочно спаренных нуклеотидов
Процесс исправления избавляет от основной массы ошибок, но не от всех. После создания новой ДНК запускается механизм репарации ошибочно спаренных нуклеотидов — удаления и замены ошибочно спаренных нуклеотидов, оставшихся в результате репликации. Исправление несоответствий между парами оснований также может включать в себя исправление небольших вставок и делеций, возникающих вследствие «соскальзывания» полимеразы с исходной цепи squared.
Как происходит восстановление неправильно спаренных нуклеотидов? Во-первых, белковый комплекс распознаёт неправильно спаренный нуклеотид и связывается с ним. Другой комплекс разрезает ДНК в области несовпадения, а ещё одна группа ферментов отщепляет некорректный нуклеотид вместе с небольшим участком вокруг него. Затем ДНК-полимераза заполняет этот пробел правильными нуклеотидами, а фермент ДНК-лигаза сшивает разрывы в цепиsquared.
Удивительно: как белки, участвующие в восстановлении ДНК, определяют, «кто прав» во время репарации ошибочно спаренных нуклеотидов? То есть, когда два основания неправильно соединены (как G (гуанин) и T (тимин) на рисунке выше), какое из этих двух оснований должно быть удалено и заменено?
У бактерий можно отличить исходную и дочернюю цепи ДНК по метилированным основаниям. На исходной цепи ДНК есть метильные (minus, start text, C, H, end text, start subscript, 3, end subscript) группы, присоединенные к некоторым из ее оснований, а у дочерней цепи таких групп еще нетcubed.
У эукариот процессы, позволяющие идентифицировать исходную цепь при устранении несоответствий, включают распознавание одноцепочечных разрывов, которые обнаруживаются только у дочерней цепи cubed.
Механизмы репарации ДНК
С ДНК может что-нибудь случиться практически в любой момент жизни клетки, а не только во время репликации. Фактически, ДНК постоянно повреждается из-за воздействия внешних факторов: ультрафиолетового излучения и радиации, химических веществ, не говоря уже о спонтанных процессах, которые протекают даже без вмешательства окружающей среды!start superscript, 4, end superscript
К счастью, наши клетки имеют механизмы восстановления, с помощью которых они находят и исправляют большинство повреждений ДНК. Можно выделить несколько типов репарации:
-
Прямая репарация. Некоторые повреждения ДНК, вызванные химическими реакциями, могут быть «исправлены» находящимися в клетке ферментами.
-
Эксцизионная репарация. Повреждение одного или нескольких нуклеотидов ДНК часто исправляется удалением и заменой поврежденного участка. При эксцизионной репарации оснований удаляется только поврежденное основание. В случае эксцизионной репарации нуклеотидов, как и в случае репарации ошибочно спаренных нуклеотидов, которое мы рассмотрели выше, удаляются целиком нуклеотиды.
-
Репарация двухцепочечных разрывов: Существуют два основных способа: негомологичное соединение концов и гомологичная рекомбинация. Они используются для восстановления двухцепочечных разрывов ДНК (когда вся хромосома разделяется на две части).
Прямая репарация
В некоторых случаях клетка может исправить повреждение ДНК, обратив вызвавшую его реакцию. Дело в том, что «повреждение ДНК» — это, как правило, присоединение к ней лишней группы в результате химической реакции.
Например, гуанин (G) может подвергаться реакции с присоединением метильной (minus, start text, C, H, end text, start subscript, 3, end subscript) группы к атому кислорода в азотистом основании. Если это не исправить, метил-содержащий гуанин будет связываться с тимином (Т), а не с цитозином (С) во время репликации ДНК. К счастью, у людей и многих других организмов есть фермент, который может удалить метильную группу, обратив реакцию, и тем самым вернуть азотистое основание в нормальное состояниеstart superscript, 5, end superscript.
Эксцизионная репарация оснований
Эксцизионная репарация оснований — это механизм, используемый для обнаружения и удаления определенных типов поврежденных азотистых оснований. Ключевую роль в нем играет группа ферментов, называемых гликозилазами. Каждая гликозилаза обнаруживает и удаляет определенный вид поврежденных оснований.
Например, в процессе реакции дезаминирования цитозин может превратиться в урацил — основание, обычно встречающееся только в РНК. Во время репликации ДНК урацил будет соединяться с аденином, а не с гуанином (в отличие от цитозина), поэтому такое превращение может привести к возникновению мутацииstart superscript, 5, end superscript.
Для предотвращения подобных изменений гликозилаза, являющаяся частью сигнального пути эксцизионной репарации, обнаруживает и удаляет дезаминированные цитозины. После того, как основание было удалено, удаляется и оставшаяся часть нуклеотида, а другие ферменты заполняют пробелstart superscript, 6, end superscript.
Эксцизионная репарация нуклеотидов
Эксцизионная репарация нуклеотидов — это еще один способ удаления и замены поврежденных оснований. В результате нее обнаруживаются и корректируются повреждения, которые искажают форму двойной спирали ДНК. Например, азотистые основания могут измениться, присоединив к себе громоздкие группы атомов, в частности, в результате воздействия химических веществ, содержащихся в сигаретном дымеstart superscript, 7, end superscript.
Эксцизионная репарация нуклеотидов также используется для устранения повреждений, вызванных ультрафиолетовым излучением, например, при получении солнечного ожога. Под воздействием УФ-излучения цитозин и тимин могут вступать в реакцию с соседними основаниями, которые также являются цитозином или тимином, образуя при этом связи, изменяющие форму двойной спирали и вызывающие ошибки в процессе репликации ДНК. Наиболее распространенный тип таких связей — тиминовый димер — он состоит из двух тиминовых оснований, вступающих в реакцию друг с другом и образующих химическую связьstart superscript, 8, end superscript.
При эксцизионной репарации нуклеотидов поврежденные нуклеотиды удаляются вместе с соседними нуклеотидами. В этом процессе хеликаза (фермент, раскручивающий ДНК) раскрывает ДНК, образуя пузырь, а ферменты, разрезающие ДНК, отсекают поврежденную часть пузыря. Полимераза заполняет пробел, а лигаза сшивает разрыв в цепиstart superscript, 9, end superscript.
Репарация двухцепочечных разрывов
Некоторые факторы окружающей среды, например, радиация, могут вызывать разрывы обеих цепочек ДНК (разделение хромосомы на две части). Такие повреждения ДНК, если верить комиксам, ведут к появлению супергероев, но могут встречаться и после реальных катастроф, например, Чернобыльской.
Двухцепочечные разрывы опасны, потому что большие сегменты хромосом и сотни содержащихся в них генов могут быть потеряны, если разрыв не будет восстановлен. Существует два способа восстановления двухцепочечных разрывов ДНК: негомологичное соединение концов и гомологичная рекомбинация.
При негомологичном соединении концов два разорванных конца хромосомы просто склеиваются обратно. Этот механизм восстановления является «грубым» и неточным, в результате в месте разрыва, как правило, либо теряются нуклеотиды, либо добавляются лишние, что может привести к мутациям. Но это в любом случае лучше потери целого фрагмента хромосомыstart superscript, 10, end superscript.
При гомологичной рекомбинации для восстановления разрыва используется фрагмент из гомологичной хромосомы, который соответствует поврежденной хромосоме (или из сестринской хроматиды, если ДНК была реплицирована). В этом процессе две хромосомы объединяются, и неповрежденная область гомологичной хромосомы или хроматиды используется в качестве матрицы для замены поврежденной области. Гомологичная рекомбинация работает «чище», точнее, чем негомологичное соединение концов, и обычно не приводит к образованию мутацийstart superscript, 11, end superscript.
Репарация ДНК и заболевания человека
Доказательства важности механизмов репарации получены на основе генетических заболеваний человека. Во многих случаях мутации в генах, которые кодируют белки, участвующие в репарации, связаны с наследственным раком. Например:
-
Наследственный неполипозный колоректальный рак (также называемый синдромом Линча) вызван мутациями в генах, кодирующих белки, которые участвуют в репарации ошибочно спаренных нуклеотидовstart superscript, 12, comma, 13, end superscript. Поскольку такие нуклеотиды не восстанавливаются, у людей, страдающих этим синдромом, мутации накапливаются гораздо быстрее, чем у здоровых. Это может привести к развитию опухолей толстой кишки.
-
Люди с пигментной ксеродермой очень чувствительны к ультрафиолетовому излучению. Это вызвано мутациями в белках, участвующих в эксцизионной репарации нуклеотидов. Когда они не функционируют, димеры тимина и другие виды повреждений, вызванные ультрафиолетовым излучением, перестают восстанавливаться. У людей с пигментной ксеродермой после нескольких минут пребывания на солнце могут возникнуть сильные солнечные ожоги, и около половины из них заболевают раком кожи в возрасте до 10 лет, если только они не избегают солнечных лучейstart superscript, 14, end superscript.
Макеты страниц
Было установлено, что частота ошибок при репликации ДНК Е. coli не превышает 1 на  нуклеотидов. Поскольку хромосома Е. coli содержит приблизительно
нуклеотидов. Поскольку хромосома Е. coli содержит приблизительно  пар оснований, на 10000 клеток, претерпевших один цикл деления, встраивается всего один неправильный нуклеотид.
пар оснований, на 10000 клеток, претерпевших один цикл деления, встраивается всего один неправильный нуклеотид.
Долгое время считалось, что столь высокая степень точности воспроизведения генетической информации целиком определяется точностью уотсон-криковского спаривания между матричной и новообразованной (дочерней) цепями, однако в результате последующего анализа выяснилось, что если бы точность репликации зависела исключительно от правильности спаривания оснований, то частота ошибок была бы значительно выше — приблизительно 1 на 104-105 остатков. Следовательно, чтобы объяснить такую низкую частоту ошибок при репликации in vivo, необходимо предположить участие в процессе репликации еще какого-то одного или нескольких факторов.
Более детальное изучение свойств высокоочищенных ДНК-полимераз позволило получить по крайней мере частичный ответ на вопрос о природе этих факторов. Напомним, что ДНК-полимеразы I и III обладают тремя различными ферментативными активностями. Мы уже видели, как фермент функционирует в качестве полимеразы, а также как он может удалять нуклеотидные остатки с 5-конца фрагмента ДНК. Однако 3-экзонуклеазная активность ДНК-полимераз I и III очень озадачивала исследователей, ибо она означала, что эти ферменты способны «пятиться», отщепляя З-концевые нуклеотиды в направлении, противоположном тому, в котором они действуют как полимеразы. З-экзону-клеазная активность ДНК-полимераз I и III — это средство проверки новосинтезированной цепи ДНК и исправления ошибок, сделанных ферментом при его работе в качестве полимеразы. Если ДНК-полимераза встраивает неправильный нуклеотид, то фермент сам может распознать неспособность этого нуклеотида образовать правильную пару с соответствующим нуклеотидом матрицы (рис. 28-15). В этом случае фермент возвращается назад и отщепляет неправильный нуклеотид с З-конца цепи, после чего полимераза продолжает присоединять правильные нуклеотиды, т.е. возобновляет свое обычное продвижение в направлении 

Рис. 28-15. Исправление ошибок с помощью 3-зкзонуклеазной активности ДНК-полимеразы.
Таким образом, по мере перемещения репликативной вилки вдоль матрицы осуществляется проверка каждого встроенного нуклеотида. Корректирующее действие ДНК-полимеразы очень эффективно; благодаря ему точность репликации повышается как минимум в 104 раз. Суммарная ошибка возникает в результате ошибок, допускаемых ферментом в ходе полимеризации и в процессе исправления их при корректировке; она не превышает одной ошибки на  нуклеотидных остатков.
нуклеотидных остатков.
Очень важно отметить, что процесс репликации протекает со значительно более высокой степенью точности, чем процессы транскрипции и трансляции. Частые ошибки в репликации подвергли бы большому риску сохранность видов и их жизнеспособность.
Ошибки же в транскрипции и трансляции гораздо менее опасны, поскольку они влияют на образование РНК или белка только в одной клетке и не изменяют всю последующую родословную вида. Корректировка с помощью ДНК-полимеразы — это, вероятно, лишь один из путей, обеспечивающих высокую точность репликации. Возможно, исключительно сложная организация репликативного процесса и участие в нем множества белков необходимы для достижения именно этой цели. Интересно, что некоторые эукариотические ДНК-полимеразы не осуществляют корректировку. По-видимому, эукариоты обеспечивают точность и надежность процесса репликации с помощью каких-то других средств.
Причины мутаций
Точечные
мутации, суть которых связана с заменами
одной пары оснований на другую пару,
могут быть вызваны самыми различными
причинами. К числу наиболее известных
и хорошо изученных факторов, вызывающих
появление точечных мутаций относятся
нижеследующие.
Ошибки
репликации, не исправленные ДНК-полимеразой.
Как известно, точность копирования в
процессе репликации ДНК настолько
велика, что в среднем на каждые 1 10 9
пар нуклеотидов приходится одна
ошибка. Такую высокую точность репликации
обеспечивает корректирующая (3′
5′)-экзонуклеазная активность
ДНК-полимеразы. Тем не менее, в ряде
случаев ДНК-полимераза может ошибаться.
Одной из причин таких ошибок является
способность всех азотистых оснований
образовывать термодинамически невыгодные
таутомерные формы за счет миграции
атома водорода. При этом амино- и
оксогруппы превращаются в иминогруппы
(=NH) и енольные группы
(-ОН), соответственно. Такие редкие
таутомерные формы, как правило, образуют
неправильные, неканонические пары с
другими основаниями. Примером может
служить способность Cyt образовывать
редкую таутомерную имино-форму,
приведенную на рис 5.4.

|
Рис. 5.4 |
Таутомерная |
Эта
имино-форма
Cyt
образует пару не с Gua, а с Ade. В результате,
в процессе последующей репликации,
может произойти замена пары A-T на G-C.
Точно также аденин способен образовывать
редкую таутомерную имино-форму, которая
приобретает способность комплементарно
спариваться с неканоническим для него
цитозином (рис. 5.5).
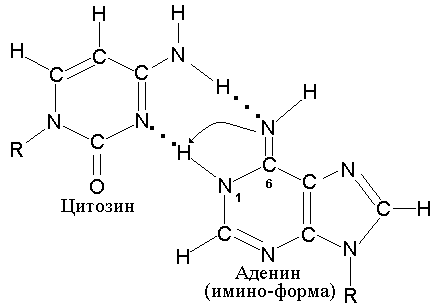
|
Рис. 5.5 |
Таутомерная |
Процессы
спонтанного дезаминирования обычных
и модифицированных оснований. В
настоящее время хорошо известно, что
некоторая часть оснований, входящих в
состав ДНК как про-, так и эукариот
метилируется пострепликативно особыми
ферментами — метилазами.
Цели метилирования у про- и эукариот
различны, но эта модификация ДНК протекает
достаточно интенсивно. Чаще всего
метилированию подвергаются остатки
Ade и Cyt. Ранее нами была рассмотрена
схема, показывающая последовательность
событий приводящих к возникновению
транзиции G-C
A-T
вследствие спонтанного или индуцированного
дезаминирования цитозина (рис. 5.1), а
также причины существования горячих
точек.
Например,
если дезаминированию подвергается не
цитозин, а 5-метилцитозин,
то этот процесс приводит к образованию
нормального для днк
основания – тимина (Thy). Естественно,
что в этом случае при повторной репликации
возможна замена пары G-C на пару A-T. Было
обнаружено, что процессы спонтанного
дезаминирования происходят с определенной
достаточно высокой скоростью, которая
составляет 100 актов дезаминирования на
один геном в сутки.
Процессы
апуринизации ДНК.
ДНК каждой клетки человеческого организма
в результате апуринизации
теряет в сутки около 5.000 пуриновых
оснований (Ade и Gua) вследствие термального
разрыва N-гликозидных связей между
пуриновым основанием и дезоксирибозой.
Удаление пиримидиновых оснований из
ДНК в какой-либо ощутимой степени не
происходит из-за того, что N-гликозидные
связи пиримидинов с углеводной частью
намного более стабильны, чем связь
пурина с углеводом. На интенсивность
процесса апуринизации ДНК могут влиять
также различные химические факторы.
Например, в кислой среде эффективность
апуринизации ДНК существенно возрастает.
Алкилирование гуанина под действием
диметилсульфата приводит к образованию
четвертичного азота в 7-ом положении
этого азотистого основания, что ослабляет
N-гликозидную связь с дезоксирибозой и
высвобождает метильное производное
гуанина, обеспечивая дополнительную
потерю пуринов молекулами ДНК (рис.
5.6).


.
|
Рис. 5.6 |
Алкилирующий |
Повреждения
ДНК, вызываемые действием химических
факторов окружающей среды.
Основания в составе ДНК весьма
чувствительны к действию многочисленных
химических соединений, распространенных
в окружающей среде. Многие из них получили
название ксенобиотиков, большинство
которых имеет антропогенное техногенное
происхождение. К разряду ксенобиотиков
относятся многочисленные яды, лекарства,
канцерогены, пестициды, инсектициды,
гербициды и многие другие соединения.
Среди эффектов различных соединений
наиболее полно изучено воздействие
азотистой кислоты (HNO2)
(рис. 5.3), гидроксиламина (NH2OH)
(рис.5.7), алкилирующих агентов, таких
как диметилсульфат (рис. 5.6),
N-метил-N/-нитро-N-нитрозогуанидин:
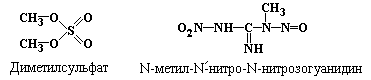

.
|
Рис. 5.7 |
Реакция цитозина |
Под
действием азотистой
кислоты,
которая может образовываться из таких
предшественников как NaNO2
— нитрит натрия, NaNO3
— нитрат натрия, а также органических
соединений типа нитрозаминов

происходит
активное дезаминирование Cyt с образованием
Ura, Ade с образованием гипоксантина (Hyp) и
Gua с образованием ксантина (Xan). В
результате дезаминирования Cyt образуется,
как уже известно, Ura, который комплементарно
спаривается с Ade. В результате происходит
транзиция G-C
A-T.
С
другой стороны, в результате дезаминирования
Ade образуется Hyp, который приобретает
способность спариваться с Cyt, вызывая
транзицию A-T
G-C. если
события, касающиеся дезаминирования,
произойдут в одном сайте, т.е. сначала
дезаминированию подвергнется Cyt и
произойдет образование A-T пары вместо
G-C, а затем дезаминированию подвергнется
появившийся Ade и произойдет обратная
транзиция A-T
G-C, то при этом последовательность ДНК
восстановится.
Дезаминирование
Gua в Xan не влияет на способность измененного
основания образовывать пару с цитозином
(Xan – Cyt).
Алкилирующие
агенты могут
воздействовать как на структуру
оснований, так могут разрушать и
фосфодиэфирные связи, приводя к
фрагментированию цепей ДНК. Кроме того,
некоторые алкилирующие агенты способны
ковалентно взаимодействовать с обеими
цепями ДНК, вызывая образование поперечных
сшивок.
Кроме
упомянутого выше диметилсульфата, к
числу наиболее активных алкилирующих
агентов относятся
диметилнитрозамин и
азотистый
иприт:
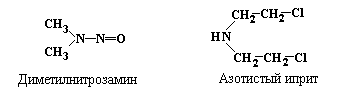
В результате
воздействия алкилирующих (как метилирующих,
так и этилирующих) агентов может
происходить метилирование Gua по
7-положению, что приводит к образованию
7-метилгуанина (рис. 5.8), который впоследствие
образует неканоническую пару с тимином.
С тимином также может спариваться
7-этилгуанин.

|
Рис. 5.8 |
Взаимодействие |
Образование
неканонической пары между тимином и
7-этилгуанином показано на рис. 5.9.

|
Рис. 5.9 |
Характер |
Кроме
приведенных выше способов модификации
гуанина, это азотистое основание может
также метилироваться по гидроксильной
группе енольной формы с образованием
O6-метил-Gua,
не способного образовывать нормальную
комплементарную пару с Cyt. В свою очередь,
метилирование аденина по аминогруппе
приводит к образованию N6-метиладенина.
Ошибки
ДНК-полимеразы,
связанные
с
включением
аналогов природных нуклеотидов.
Одним из аналогов природных нуклеотидов,
не выщепляемых ДНК-полимеразой является
нуклеотид с азотистым основанием —
2-аминопурином (рис. 5.10), который
встраивается в ДНК вместо Ade, но
впоследствии спаривается с цитозином
и тем самым способствует транзиции A-T
G-C.

|
Рис. 5.10 |
Аналог аденина |
Несмотря
на высокую точность функционирования
ДНК-полимераз при катализе репликации
эти ферменты не всегда способны отличать
нормальные дезоксирибонуклеозид
трифосфатные субстраты от некоторых
других нуклеотидов с очень похожей
структурой. Следует отметить, что в
случае 2-аминопурина ДНК-полимераза все
же делает существенную ошибку, поскольку
наличие NH2-группы
во 2-ом или в 6-ом положениях пурина
структурно является очень заметным. На
рис. 5.11, приведенном ниже, представлена
схема возникновения транзиции A-T
G-C, инициируемая встраиванием 2-аминопурина.

|
Рис. 5.11 |
Схема, отражающая |
Другой
пример таких структурных аналогов
природных нуклеозидтрифосфатов –
5-Br-dUTP
(5-Br-дезоксиуридинтрифосфат),
который является аналогом тимидинтрифосфата
из-за присутствия атома Br в 5-положении
урацила, где у тимина находится CH3-группа.
Первоначально 5-галоидпроизводные
урацила были синтезированы как аналоги
тимина с целью их возможного применения
в качестве цитостатиков или противовирусных
средств. В частности цитостатические
эффекты 5-Br-dUrd сводятся к эффективному
фосфорилированию данного аналога
тимидина под действием тимидинкиназы
поврежденной клетки и встраиванию этого
модифицированного нуклеотида в ДНК. В
результате последующего облучения
клеток ультрафиолетовым светом встроенный
5-Br-Ura принимает участие в образовании
большого числа пиримидиновых димеров
и поперечных сшивок в ДНК, что практически
полностью блокирует возможность
репликации или транскрипции ДНК. Ситуация
с 5-Br–dUrd, как соединением способным
инициировать транзиции, менее однозначна,
чем в случае 2-аминопурина. Дело в том,
что 5-Br-dUrd может образовывать кето- и
енольную формы. Минорная енольная форма
5-Br-Ura (рис. 5.12) возникает чаще, чем такая
же форма тимина, из-за большей
электроотрицательности Br по сравнению
CH3-группой
тимина.

|
Рис. 5.12 |
Высокая |
Поэтому,
более часто образуемая енольная форма
5-Br-Ura имеет тенденцию спариваться с
Gua, что приводит к транзиции A-T
G-C. в
норме же, кето-форма 5-Br-Ura, являющегося
аналогом тимина спаривается с Ade (рис.
5.13).
|
Рис. 5.13 |
В норме кето-форма |
Схема,
иллюстрирующая последовательность
этапов приводящих к транзиции A-T
G-C, вызванной енолизацией встроенного
в ДНК 5-Br-Ura показана на рис. 5.14.

|
Рис. 5.14 |
Схема появления |
Явление
интеркаляции.
Некоторые органические соединения,
которые характеризуются плоской
ароматической структурой с соответствующей
геометрией и размерами могут встраиваться
в ДНК между парами оснований —
интеркалировать.
В результате интеркаляции, эти соединения
вызывают появление вставок или делеций
одной или более пар оснований и тем
самым приводят к изменению рамки
считывания, если только вставки и
делеции не кратны трем парам оснований.
К таким интеркалирующим соединениям
относятся акридины
и этидий
бромид (рис.
5.15).

|
Рис. 5.15 |
Этидий бромид |
Рентгеноструктурный
анализ комплексов таких соединений с
синтетическими двухцепочечными
олигонуклеотидами показывает, что
плоские ароматические кольца акридиновых
красителей
внедряются между парами оснований
двойной спирали. Механизм внедрения
предполагает проникновение молекулы
красителя между парами оснований в
момент возникновения локального
нарушения структуры, при этом водородные
связи между парами оснований сохраняются,
тогда как «стэкинг»- взаимодействия
нарушаются.
Одним
из вариантов такого нарушения структуры
ДНК вследствие интеркаляции акридиновых
красителей
или
этидий
бромида
является образование изломов двойной
спирали в молекуле ДНК, которые получили
название кинков.
Химический
канцерогенез.
В настоящее время многие эксперты
считают, что в подавляющем большинстве
случаев заболевание раком инициируется
воздействием на нуклеиновые кислоты
определенных химических соединений.
Канцерогенные вещества поступают в
окружающую среду не только благодаря
синтезу и использованию новых химических
соединений в промышленных масштабах.
Канцерогенами являются также многие
соединения естественного происхождения.
Например, известными канцерогенами
являются афлатоксины,
продуцируемые некоторыми плесневыми
грибками. Несмотря на то, что интенсивное
изучение микотоксинов
началось сравнительно недавно, к
настоящему времени уже описано более
300 таких соединений, относящихся к 25
различным типам. Даже в небольших дозах
микотоксины оказывают разнообразные
токсические эффекты на человека и
животных, приводят к деградации печени,
геморрагии и карциноме. В качестве
главных по опасности микотоксинов
сейчас рассматривают группу метаболитов
гриба Aspergillus
flavus
– афлатоксинов, из которых наиболее
коварны афлатоксин
В1
и продукт его метаболического
гидроксилирования в организме коровы,
проникающий в молоко – афлатоксин
М1
(рис. 5.16). Доказано, что эти соединения
являются причиной цирроза и рака печени
у людей. Механизм действия афлатоксинов
состоит в том, что они после
15,16-эпоксидирования с участием печеночного
цитохрома Р-450 ковалентно связываются
с РНК, блокируя синтез белка.
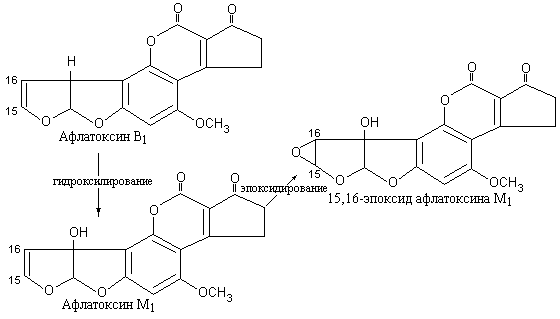
|
Рис. 5.16 |
Метаболическая |
К
другой группе канцерогенов, имеющих
естественное происхождение, относятся
такие соединения, как бензпирен
и бензантрацен,
являющиеся постоянными компонентами
табачного дыма, а также копченых
продуктов питания и продуктов
приготовляемых на углях. Хорошо известно,
что некоторые канцерогены характеризуются
непосредственным воздействием на
нуклеиновые кислоты, тогда как другие
(бензпирен и бензантрацен сами по себе
являются слабыми канцерогенами) прежде,
чем стать канцерогенными, должны пройти
стадию активации посредством
гидроксилирования и эпоксидирования
с участием монооксигеназных систем
печени. Ферменты, которые катализируют
активацию канцерогенов, принадлежат к
семейству цитохромов P-450. Как было
показано, конечным продуктом активации
бензпирена является вещество обладающее
мощным канцерогенным действием на
человека и животных и сильнейшим
мутагенным эффектом на бактериальные
клетки. Это соединение представляет
собой 7,8-дигидродиол-9,10-эпоксид
бензпирена.
Схема реакций, приводящих к метаболической
активации бензпирена приведена на рис.
5.17.
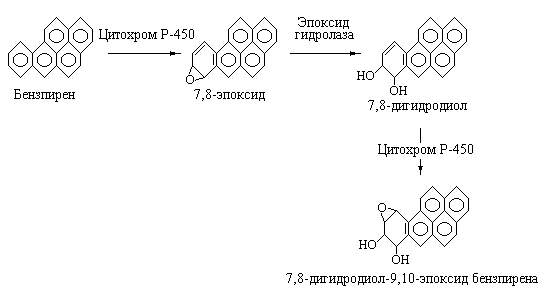
|
Рис. 5.17 |
Метаболическая |
На
первой стадии в описываемой
последовательности реакций бензпирен
под действием цитохрома Р-450 превращается
в 7,8-эпоксид, который далее с участием
эпоксидгидролазы присоединяет воду с
образованием 7,8-дигидродиола бензпирена.
На последней стадии, катализируемой
также цитохромом Р-450 образуется конечное
соединение 7,8-дигидродиол-9,10-эпоксид
бензпирена.
Следует
иметь в виду, что цитохромы P-450 обладают
уникальной способностью индуцироваться
неканцерогенными соединениями, такими
как этанол. Следовательно, алкоголь
может в значительной степени увеличивать
потенциальный риск рака в результате
воздействия канцерогенов.
Мутагенное
действие физических факторов
(X-лучи, УФ-,
-излучение).
Как ультрафиолетовое, так и рентгеновское
излучения являются сильнейшими
мутагенными средствами. Нормальные
основания, входящие в состав ДНК,
представлены, как известно, в виде кето-
и амино-форм,
находящихся в равновесии с очень
небольшими количествами минорных
енольной
и имино-форм.
Энергия УФ и X-лучей сдвигает это
равновесие в сторону образования
минорных таутомерных форм. В результате
чего повышенное содержание редких
таутомерных форм Ade и Cyt увеличивает
частоту их спаривания с Cyt и Ade,
соответственно (рис. 5.4 и 5.5). Считают,
что повышенное количество енольных
форм оснований в момент репликации
значительно повышает частоту мутаций
в новосинтезируемых цепях ДНК.
Действие
на ДНК жесткого излучения типа
рентгеновских лучей и -излучения
может приводить к изменению структуры
оснований. Результатом такого воздействия
может быть раскрытие гетероциклов,
разрушение фосфодиэфирных связей. В
присутствии кислорода накапливается
большое количество продуктов окисления
азотистых оснований и остатков
дезоксирибозы.
При
воздействии УФ-света, кроме сдвига
равновесия в сторону образования
минорных таутомерных форм оснований
происходит также появление, в первую
очередь, тиминовых димеров (рис. 5.18),
хотя в принципе возможно образование
ковалентно сшитых пар не только T-T, но
также T-C и C-C.

|
Рис. 5.18 |
Структура |
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Удвоение генетического материала перед делением клетки — очень точный процесс. Но мутации в геноме не перестают накапливаться, что приводит как к болезням, так и к появлению нового материала для эволюции. Оказывается, одним из механизмов сохранения мутаций может служить связывание белков — полимераз и транскрипционных факторов — с ДНК. Они создают помеху для «выщепления» ошибочных последовательностей, синтезированных неточной ДНК-полимеразой α.

Рисунок 1. Схема репликативной вилки. Новая цепь синтезируется на матрице старой в направлении от 5’- к 3’-концу. При этом одна из цепей может расти непрерывно, а другая — нет (очередное праймирование происходит по мере расплетения ДНК). Рисунок с сайта topreferat.znate.ru.
Перед тем как поделиться, клетке нужно дублировать свой генетический материал. Процесс удвоения молекул ДНК называется репликацией. Репликация осуществляется и регулируется многими белками. Она должна быть максимально точной, чтобы избежать появления мутаций в ДНК. Механизм удвоения генетического материала у прокариот и эукариот полуконсервативный. То есть после репликации одной двуцепочечной молекулы ДНК получаются две, в каждой из которых одна цепь будет старой (материнской), а другая новой (дочерней) [1]. Образовавшиеся молекулы ДНК почти идентичные. Но их минорные отличия могут играть важную роль в судьбе организма [2, 3].
Немного о ДНК-полимеразах
Ключевыми белками репликации являются ДНК-полимеразы — ферменты, читающие цепочку нуклеотидов и использующие ее как матрицу для синтеза дочерней молекулы ДНК. Важно отметить, что ДНК-полимеразы могут присоединять новый нуклеотид только к предыдущему, потому им всегда нужна «затравка» (праймер). Такие затравки во время репликации синтезирует другой фермент — праймаза. Издержки такого разделения труда заключаются в том, что первым компонентом дочерней цепи становятся рибонуклеотиды, то есть короткие цепочки РНК. К ним полимераза уже может присоединить дезоксирибонуклеотиды. Это делает ДНК-полимераза α. Она удлиняет РНК-затравку, синтезируя небольшую цепочку ДНК (примерно 20 нуклеотидов). Так как репликация должна быть очень точной, то неудивительно, что некоторые ДНК-полимеразы умеют исправлять ошибки своей работы. В первую очередь это касается ферментов, синтезирующих длинные участки цепей. Полимераза α такой редактирующей способностью не обладает, поэтому включает новые нуклеотиды, не «оглядываясь» назад и оставляя много ошибок.
Каждая цепь ДНК имеет направление: у нее есть 3’- и 5’-концы. Важно отметить, что новая молекула ДНК может удлиняться только в одну сторону, так как ДНК-полимеразы умеют добавлять нуклеотиды только к 3’-концу. В начале репликации две цепи материнской ДНК расплетаются в определенном месте, и на их матрицах строятся дочерние цепи в двух направлениях. В одну сторону синтез идет непрерывно, а в другую — с помощью коротких фрагментов, как показано на рисунке 1. Такой механизм обеспечивает удлинение дочерних цепей только от 5’-конца к 3’-концу [4, 5]. Цепь, синтез которой идет короткими фрагментами — фрагментами Оказаки, называется отстающей.
У эукариот синтез всего фрагмента Оказаки от уже продленной полимеразой α затравки осуществляет полимераза δ. Она очень точная и умеет исправлять свои ошибки. Получается, что в начале каждого фрагмента Оказаки есть небольшой фрагмент РНК и участок, синтезированный «малограмотной» полимеразой α. Всё это нужно удалить и застроить заново полимеразой δ, так как ни РНК, ни ошибок в дочерней цепи быть не должно (рис. 2). Поле такого редактирования фрагменты Оказаки сшиваются ДНК-лигазой, образуя непрерывную цепь [6, 7]. Но, несмотря на исправление ошибок, некоторые из них все-таки остаются, и возникают мутации.

Рисунок 2. Структура фрагмента Оказаки. В начале фрагмента есть часть, которая особенно нуждается в редактировании (ограничена пунктиром). В клетке для этого имеются специальные механизмы. Но если с ДНК быстро свяжется белок, то ошибки в 5’-концевых участках фрагментов Оказаки могут остаться неисправленными.
Возможный механизм сохранения мутаций после репликации
Показано, что мутации в геноме происходят неравномерно [8]. Их распределение зависит от многих факторов, в том числе и от деятельности ДНК-связывающих белков. Авторы недавнего исследования сопоставили распределение мутаций в геноме и участков «стыка» фрагментов Оказаки (рис. 3а). Получилось, что на центр нуклеосомной ДНК приходится как пик мутаций, так и максимальное количество концов фрагментов Оказаки [9].
Такая корреляция была обнаружена и в зонах связывания транскрипционных факторов (рис. 3b). Правда, непосредственно специфические сайты посадки белков мутируют редко. Это, видимо, связано с давлением отбора: если сайт связывания транскрипционного фактора будет поврежден, то может сбиться регуляция важных процессов, что в свою очередь с высокой вероятностью приведет к гибели клетки.

Рисунок 3. Корреляция нуклеотидного полиморфизма (числа мутаций) и количества стыков фрагментов Оказаки в местах посадки белков (нуклеосомных — слева, транскрипционных регуляторов — справа) на ДНК дрожжей S. cerevisiae. Рисунок из [9].
Синтез фрагментов Оказаки имеет направление. Если мутации связаны с этим процессом, то их распределение тоже должно иметь направление. Выявили, что количество мутаций резко возрастает сразу после окончания фрагмента Оказаки. Особенно в местах связывания белков, служащих барьером для полимеразы δ [10].
Авторы показали, что совпадение мест стыка фрагментов Оказаки и повышенного уровня мутаций является следствием именно того, что в этих участках с ДНК связываются белки. Последовательности ДНК в таких регионах всегда разные, поэтому не имеют прямого отношения к наблюдаемой закономерности.
Как было сказано выше, фрагменты Оказаки синтезируются полимеразами α и δ. Ранее полагали, что в ходе этого процесса большинство нуклеотидов (если не все), синтезированных полимеразой α, удаляются [11, 12]. Это было бы полезно, потому что у полимеразы α нет возможности исправления ошибок. Но авторы исследования предполагают, что «следы» полимеразы α будут оставаться, если с ДНК быстро свяжется белок (рис. 4).

Рисунок 4. Модель закрепления ошибок репликации из-за белка, связанного с началом фрагмента Оказаки. Рисунок из [9].
В подтверждение этой гипотезы ученые показали, что ДНК, синтезированная полимеразой α, остается в геноме и после завершения репликации, а не исключается целиком. Мало того, она составляет 1,5% генома.
Интересно, что найденные закономерности характерны не только для дрожжей, но и для человека. Часто вокруг типичных сайтов связывания белков в ДНК накапливаются мутации. Авторы исследования связывают это именно с активностью полимеразы α.
- Meselson M., Stahl F.W. (1958). The replication of DNA in Esherichia coli. Proc. Natl. Acad. Sci. USA. 44 (7), 671–682;
- Генная терапия против рака;
- Союз голубоглазых;
- Johnston L.H., Nasmyth K.A. (1978). Saccharomyces cerevisiae cell cycle mutant cdc9 is defective in DNA ligase. Nature. 274, 891–893;
- Okazaki R., Okazaki T., Sakabe K., Sugimoto K., Sugino A. (1968). Mechanism of DNA chain growth. I. Possible discontinuity and unusual secondary structure of newly synthesized chains. Proc. Natl. Acad. Sci. USA. 59, 598–605;
- Balakrishnan L. and Bambara R.A. (2013). Okazaki fragment metabolism. Cold Spring Harb. Perspect. Biol. 5 (2), doi: 10.1101/cshperspect.a010173;
- Zheng L. and Shen B. (2011). Okazaki fragment maturation: nucleases take centre stage. J. Mol. Cell Biol. 3, 23–30;
- Wolfe K.H., Sharp P.M., Li W.H. (1989). Mutation rates differ among regions of the mammalian genome. Nature. 337, 283–285;
- Reijns M.A.M., Kemp H., Ding J., de Proce S.M., Jackson A.P., Taylor M.S. (2015). Lagging-strand replication shapes the mutational landscape of the genome. Nature. 518, 502–506;
- Smith D.J. and Whitehouse I. (2012). Intrinsic coupling of lagging-strand synthesis to chromatinassembly. Nature. 483, 434–438;
- Perera R.L., Torella R., Klinge S., Kilkenny M.L., Maman J.D., Pellegrini L. (2013). Mechanism for priming DNA synthesis by yeast DNA Polymerase alpha. eLife 2013;2:e00482;
- Walsh E. and Eckert K.A. Eukaryotic replicative DNA Polymerases. In: Murakami K. and Trakselis M.A. (Eds.), Nucleic Acid Polymerases. Springer-Verlag Berlin Heidelberg, 2014. 30, 17–41. ISBN 978-3-642-39795-0..