Причины мутаций
Точечные
мутации, суть которых связана с заменами
одной пары оснований на другую пару,
могут быть вызваны самыми различными
причинами. К числу наиболее известных
и хорошо изученных факторов, вызывающих
появление точечных мутаций относятся
нижеследующие.
Ошибки
репликации, не исправленные ДНК-полимеразой.
Как известно, точность копирования в
процессе репликации ДНК настолько
велика, что в среднем на каждые 1 10 9
пар нуклеотидов приходится одна
ошибка. Такую высокую точность репликации
обеспечивает корректирующая (3′
5′)-экзонуклеазная активность
ДНК-полимеразы. Тем не менее, в ряде
случаев ДНК-полимераза может ошибаться.
Одной из причин таких ошибок является
способность всех азотистых оснований
образовывать термодинамически невыгодные
таутомерные формы за счет миграции
атома водорода. При этом амино- и
оксогруппы превращаются в иминогруппы
(=NH) и енольные группы
(-ОН), соответственно. Такие редкие
таутомерные формы, как правило, образуют
неправильные, неканонические пары с
другими основаниями. Примером может
служить способность Cyt образовывать
редкую таутомерную имино-форму,
приведенную на рис 5.4.

|
Рис. 5.4 |
Таутомерная |
Эта
имино-форма
Cyt
образует пару не с Gua, а с Ade. В результате,
в процессе последующей репликации,
может произойти замена пары A-T на G-C.
Точно также аденин способен образовывать
редкую таутомерную имино-форму, которая
приобретает способность комплементарно
спариваться с неканоническим для него
цитозином (рис. 5.5).
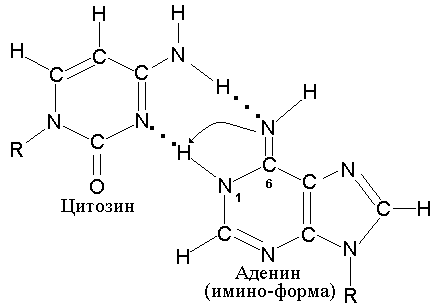
|
Рис. 5.5 |
Таутомерная |
Процессы
спонтанного дезаминирования обычных
и модифицированных оснований. В
настоящее время хорошо известно, что
некоторая часть оснований, входящих в
состав ДНК как про-, так и эукариот
метилируется пострепликативно особыми
ферментами — метилазами.
Цели метилирования у про- и эукариот
различны, но эта модификация ДНК протекает
достаточно интенсивно. Чаще всего
метилированию подвергаются остатки
Ade и Cyt. Ранее нами была рассмотрена
схема, показывающая последовательность
событий приводящих к возникновению
транзиции G-C
A-T
вследствие спонтанного или индуцированного
дезаминирования цитозина (рис. 5.1), а
также причины существования горячих
точек.
Например,
если дезаминированию подвергается не
цитозин, а 5-метилцитозин,
то этот процесс приводит к образованию
нормального для днк
основания – тимина (Thy). Естественно,
что в этом случае при повторной репликации
возможна замена пары G-C на пару A-T. Было
обнаружено, что процессы спонтанного
дезаминирования происходят с определенной
достаточно высокой скоростью, которая
составляет 100 актов дезаминирования на
один геном в сутки.
Процессы
апуринизации ДНК.
ДНК каждой клетки человеческого организма
в результате апуринизации
теряет в сутки около 5.000 пуриновых
оснований (Ade и Gua) вследствие термального
разрыва N-гликозидных связей между
пуриновым основанием и дезоксирибозой.
Удаление пиримидиновых оснований из
ДНК в какой-либо ощутимой степени не
происходит из-за того, что N-гликозидные
связи пиримидинов с углеводной частью
намного более стабильны, чем связь
пурина с углеводом. На интенсивность
процесса апуринизации ДНК могут влиять
также различные химические факторы.
Например, в кислой среде эффективность
апуринизации ДНК существенно возрастает.
Алкилирование гуанина под действием
диметилсульфата приводит к образованию
четвертичного азота в 7-ом положении
этого азотистого основания, что ослабляет
N-гликозидную связь с дезоксирибозой и
высвобождает метильное производное
гуанина, обеспечивая дополнительную
потерю пуринов молекулами ДНК (рис.
5.6).


.
|
Рис. 5.6 |
Алкилирующий |
Повреждения
ДНК, вызываемые действием химических
факторов окружающей среды.
Основания в составе ДНК весьма
чувствительны к действию многочисленных
химических соединений, распространенных
в окружающей среде. Многие из них получили
название ксенобиотиков, большинство
которых имеет антропогенное техногенное
происхождение. К разряду ксенобиотиков
относятся многочисленные яды, лекарства,
канцерогены, пестициды, инсектициды,
гербициды и многие другие соединения.
Среди эффектов различных соединений
наиболее полно изучено воздействие
азотистой кислоты (HNO2)
(рис. 5.3), гидроксиламина (NH2OH)
(рис.5.7), алкилирующих агентов, таких
как диметилсульфат (рис. 5.6),
N-метил-N/-нитро-N-нитрозогуанидин:
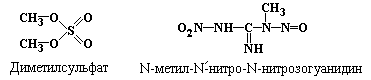

.
|
Рис. 5.7 |
Реакция цитозина |
Под
действием азотистой
кислоты,
которая может образовываться из таких
предшественников как NaNO2
— нитрит натрия, NaNO3
— нитрат натрия, а также органических
соединений типа нитрозаминов

происходит
активное дезаминирование Cyt с образованием
Ura, Ade с образованием гипоксантина (Hyp) и
Gua с образованием ксантина (Xan). В
результате дезаминирования Cyt образуется,
как уже известно, Ura, который комплементарно
спаривается с Ade. В результате происходит
транзиция G-C
A-T.
С
другой стороны, в результате дезаминирования
Ade образуется Hyp, который приобретает
способность спариваться с Cyt, вызывая
транзицию A-T
G-C. если
события, касающиеся дезаминирования,
произойдут в одном сайте, т.е. сначала
дезаминированию подвергнется Cyt и
произойдет образование A-T пары вместо
G-C, а затем дезаминированию подвергнется
появившийся Ade и произойдет обратная
транзиция A-T
G-C, то при этом последовательность ДНК
восстановится.
Дезаминирование
Gua в Xan не влияет на способность измененного
основания образовывать пару с цитозином
(Xan – Cyt).
Алкилирующие
агенты могут
воздействовать как на структуру
оснований, так могут разрушать и
фосфодиэфирные связи, приводя к
фрагментированию цепей ДНК. Кроме того,
некоторые алкилирующие агенты способны
ковалентно взаимодействовать с обеими
цепями ДНК, вызывая образование поперечных
сшивок.
Кроме
упомянутого выше диметилсульфата, к
числу наиболее активных алкилирующих
агентов относятся
диметилнитрозамин и
азотистый
иприт:
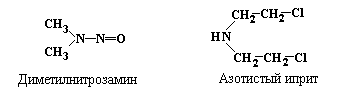
В результате
воздействия алкилирующих (как метилирующих,
так и этилирующих) агентов может
происходить метилирование Gua по
7-положению, что приводит к образованию
7-метилгуанина (рис. 5.8), который впоследствие
образует неканоническую пару с тимином.
С тимином также может спариваться
7-этилгуанин.

|
Рис. 5.8 |
Взаимодействие |
Образование
неканонической пары между тимином и
7-этилгуанином показано на рис. 5.9.

|
Рис. 5.9 |
Характер |
Кроме
приведенных выше способов модификации
гуанина, это азотистое основание может
также метилироваться по гидроксильной
группе енольной формы с образованием
O6-метил-Gua,
не способного образовывать нормальную
комплементарную пару с Cyt. В свою очередь,
метилирование аденина по аминогруппе
приводит к образованию N6-метиладенина.
Ошибки
ДНК-полимеразы,
связанные
с
включением
аналогов природных нуклеотидов.
Одним из аналогов природных нуклеотидов,
не выщепляемых ДНК-полимеразой является
нуклеотид с азотистым основанием —
2-аминопурином (рис. 5.10), который
встраивается в ДНК вместо Ade, но
впоследствии спаривается с цитозином
и тем самым способствует транзиции A-T
G-C.

|
Рис. 5.10 |
Аналог аденина |
Несмотря
на высокую точность функционирования
ДНК-полимераз при катализе репликации
эти ферменты не всегда способны отличать
нормальные дезоксирибонуклеозид
трифосфатные субстраты от некоторых
других нуклеотидов с очень похожей
структурой. Следует отметить, что в
случае 2-аминопурина ДНК-полимераза все
же делает существенную ошибку, поскольку
наличие NH2-группы
во 2-ом или в 6-ом положениях пурина
структурно является очень заметным. На
рис. 5.11, приведенном ниже, представлена
схема возникновения транзиции A-T
G-C, инициируемая встраиванием 2-аминопурина.

|
Рис. 5.11 |
Схема, отражающая |
Другой
пример таких структурных аналогов
природных нуклеозидтрифосфатов –
5-Br-dUTP
(5-Br-дезоксиуридинтрифосфат),
который является аналогом тимидинтрифосфата
из-за присутствия атома Br в 5-положении
урацила, где у тимина находится CH3-группа.
Первоначально 5-галоидпроизводные
урацила были синтезированы как аналоги
тимина с целью их возможного применения
в качестве цитостатиков или противовирусных
средств. В частности цитостатические
эффекты 5-Br-dUrd сводятся к эффективному
фосфорилированию данного аналога
тимидина под действием тимидинкиназы
поврежденной клетки и встраиванию этого
модифицированного нуклеотида в ДНК. В
результате последующего облучения
клеток ультрафиолетовым светом встроенный
5-Br-Ura принимает участие в образовании
большого числа пиримидиновых димеров
и поперечных сшивок в ДНК, что практически
полностью блокирует возможность
репликации или транскрипции ДНК. Ситуация
с 5-Br–dUrd, как соединением способным
инициировать транзиции, менее однозначна,
чем в случае 2-аминопурина. Дело в том,
что 5-Br-dUrd может образовывать кето- и
енольную формы. Минорная енольная форма
5-Br-Ura (рис. 5.12) возникает чаще, чем такая
же форма тимина, из-за большей
электроотрицательности Br по сравнению
CH3-группой
тимина.

|
Рис. 5.12 |
Высокая |
Поэтому,
более часто образуемая енольная форма
5-Br-Ura имеет тенденцию спариваться с
Gua, что приводит к транзиции A-T
G-C. в
норме же, кето-форма 5-Br-Ura, являющегося
аналогом тимина спаривается с Ade (рис.
5.13).
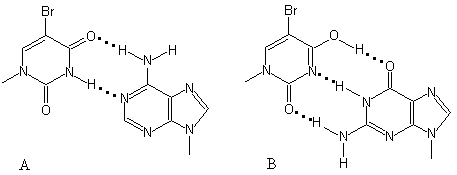
|
Рис. 5.13 |
В норме кето-форма |
Схема,
иллюстрирующая последовательность
этапов приводящих к транзиции A-T
G-C, вызванной енолизацией встроенного
в ДНК 5-Br-Ura показана на рис. 5.14.

|
Рис. 5.14 |
Схема появления |
Явление
интеркаляции.
Некоторые органические соединения,
которые характеризуются плоской
ароматической структурой с соответствующей
геометрией и размерами могут встраиваться
в ДНК между парами оснований —
интеркалировать.
В результате интеркаляции, эти соединения
вызывают появление вставок или делеций
одной или более пар оснований и тем
самым приводят к изменению рамки
считывания, если только вставки и
делеции не кратны трем парам оснований.
К таким интеркалирующим соединениям
относятся акридины
и этидий
бромид (рис.
5.15).

|
Рис. 5.15 |
Этидий бромид |
Рентгеноструктурный
анализ комплексов таких соединений с
синтетическими двухцепочечными
олигонуклеотидами показывает, что
плоские ароматические кольца акридиновых
красителей
внедряются между парами оснований
двойной спирали. Механизм внедрения
предполагает проникновение молекулы
красителя между парами оснований в
момент возникновения локального
нарушения структуры, при этом водородные
связи между парами оснований сохраняются,
тогда как «стэкинг»- взаимодействия
нарушаются.
Одним
из вариантов такого нарушения структуры
ДНК вследствие интеркаляции акридиновых
красителей
или
этидий
бромида
является образование изломов двойной
спирали в молекуле ДНК, которые получили
название кинков.
Химический
канцерогенез.
В настоящее время многие эксперты
считают, что в подавляющем большинстве
случаев заболевание раком инициируется
воздействием на нуклеиновые кислоты
определенных химических соединений.
Канцерогенные вещества поступают в
окружающую среду не только благодаря
синтезу и использованию новых химических
соединений в промышленных масштабах.
Канцерогенами являются также многие
соединения естественного происхождения.
Например, известными канцерогенами
являются афлатоксины,
продуцируемые некоторыми плесневыми
грибками. Несмотря на то, что интенсивное
изучение микотоксинов
началось сравнительно недавно, к
настоящему времени уже описано более
300 таких соединений, относящихся к 25
различным типам. Даже в небольших дозах
микотоксины оказывают разнообразные
токсические эффекты на человека и
животных, приводят к деградации печени,
геморрагии и карциноме. В качестве
главных по опасности микотоксинов
сейчас рассматривают группу метаболитов
гриба Aspergillus
flavus
– афлатоксинов, из которых наиболее
коварны афлатоксин
В1
и продукт его метаболического
гидроксилирования в организме коровы,
проникающий в молоко – афлатоксин
М1
(рис. 5.16). Доказано, что эти соединения
являются причиной цирроза и рака печени
у людей. Механизм действия афлатоксинов
состоит в том, что они после
15,16-эпоксидирования с участием печеночного
цитохрома Р-450 ковалентно связываются
с РНК, блокируя синтез белка.
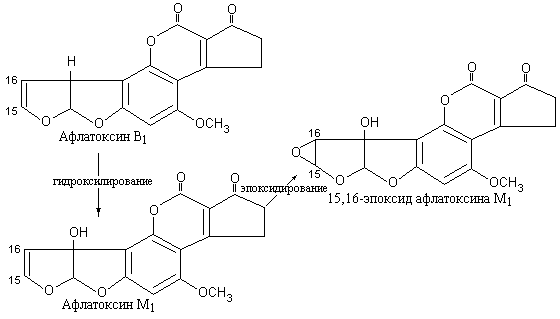
|
Рис. 5.16 |
Метаболическая |
К
другой группе канцерогенов, имеющих
естественное происхождение, относятся
такие соединения, как бензпирен
и бензантрацен,
являющиеся постоянными компонентами
табачного дыма, а также копченых
продуктов питания и продуктов
приготовляемых на углях. Хорошо известно,
что некоторые канцерогены характеризуются
непосредственным воздействием на
нуклеиновые кислоты, тогда как другие
(бензпирен и бензантрацен сами по себе
являются слабыми канцерогенами) прежде,
чем стать канцерогенными, должны пройти
стадию активации посредством
гидроксилирования и эпоксидирования
с участием монооксигеназных систем
печени. Ферменты, которые катализируют
активацию канцерогенов, принадлежат к
семейству цитохромов P-450. Как было
показано, конечным продуктом активации
бензпирена является вещество обладающее
мощным канцерогенным действием на
человека и животных и сильнейшим
мутагенным эффектом на бактериальные
клетки. Это соединение представляет
собой 7,8-дигидродиол-9,10-эпоксид
бензпирена.
Схема реакций, приводящих к метаболической
активации бензпирена приведена на рис.
5.17.
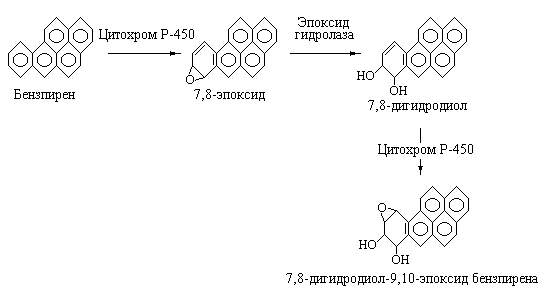
|
Рис. 5.17 |
Метаболическая |
На
первой стадии в описываемой
последовательности реакций бензпирен
под действием цитохрома Р-450 превращается
в 7,8-эпоксид, который далее с участием
эпоксидгидролазы присоединяет воду с
образованием 7,8-дигидродиола бензпирена.
На последней стадии, катализируемой
также цитохромом Р-450 образуется конечное
соединение 7,8-дигидродиол-9,10-эпоксид
бензпирена.
Следует
иметь в виду, что цитохромы P-450 обладают
уникальной способностью индуцироваться
неканцерогенными соединениями, такими
как этанол. Следовательно, алкоголь
может в значительной степени увеличивать
потенциальный риск рака в результате
воздействия канцерогенов.
Мутагенное
действие физических факторов
(X-лучи, УФ-,
-излучение).
Как ультрафиолетовое, так и рентгеновское
излучения являются сильнейшими
мутагенными средствами. Нормальные
основания, входящие в состав ДНК,
представлены, как известно, в виде кето-
и амино-форм,
находящихся в равновесии с очень
небольшими количествами минорных
енольной
и имино-форм.
Энергия УФ и X-лучей сдвигает это
равновесие в сторону образования
минорных таутомерных форм. В результате
чего повышенное содержание редких
таутомерных форм Ade и Cyt увеличивает
частоту их спаривания с Cyt и Ade,
соответственно (рис. 5.4 и 5.5). Считают,
что повышенное количество енольных
форм оснований в момент репликации
значительно повышает частоту мутаций
в новосинтезируемых цепях ДНК.
Действие
на ДНК жесткого излучения типа
рентгеновских лучей и -излучения
может приводить к изменению структуры
оснований. Результатом такого воздействия
может быть раскрытие гетероциклов,
разрушение фосфодиэфирных связей. В
присутствии кислорода накапливается
большое количество продуктов окисления
азотистых оснований и остатков
дезоксирибозы.
При
воздействии УФ-света, кроме сдвига
равновесия в сторону образования
минорных таутомерных форм оснований
происходит также появление, в первую
очередь, тиминовых димеров (рис. 5.18),
хотя в принципе возможно образование
ковалентно сшитых пар не только T-T, но
также T-C и C-C.

|
Рис. 5.18 |
Структура |
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Мутагены — факторы окружающей среды, которые способны вызывать ошибки при нормальной репликации ДНК, что ведет к спонтанным мутациям. Ошибки в репликации ДНК зависят от температурных условий, pH, состава среды. Особенно мощными мутагенными факторами являются ультрафиолетовые и ионизирующие излучения. К мутагенам химической природы относятся аналоги азотистых оснований, отдельные красители акридинового ряда, алкилирующие соединения, некоторж антибиотики, гидроксиламин, уретан и азотистая кислота. [c.61]
Химическое изменение оснований. Некоторые мутагенные вещества действуют путем химического изменения содержащихся в ДНК оснований, что приводит к ошибкам репликации. Вполне понятное изменение вызывает нитрит. Азотистая кислота дезаминирует аденин, гуанин или цитозин без разрыва или каких-либо других изменений полинуклеотидной цепи. В результате замещения аминогруппы гидроксильной группой аденин превращается в гипоксантин и спаривается с цитозином вместо тимина, что приводит к мутации АТ СС. Если цитозин дезаминируется в урацил, то он спаривается с аденином вместо гуанина, и это ведет к мутации СС -АТ. Будучи превращен в ксантин, гуанин по-прежнему спаривается с цитозином, т. е. дезаминирование С не вызывает мутации. Гидроксиламин вступает в реакцию главным образом с цитозином и изменяет его так, что тот спаривается с аденином значит, он тоже вызывает мутации СС ТА. [c.444]
Ошибки репликации ДНК приводят к мутациям [12] [c.128]
Мутации как ошибки репликации ДНК. Данные, полученные на человеке, свидетельствуют о существовании тесной связи между мутациями и клеточными делениями. Импульсом к изучению этой проблемы послужила гипотеза о механизме возникновения точковых мутаций Уотсона и Крика (рис. 5.26) [1347]. Важную роль в этом сыграли и ранние исследования на микроорганизмах, из которых следовало, что многие спонтанные мутации действительно возникают во время репликации ДНК в результате ошибочной вставки неправильного нуклеотида, приводящей к появлению в будущих клеточных поколениях новой, отличающейся пары оснований. В последние десятилетия получено до удивления мало новых данных о механизмах возникновения спонтанных мутаций, что очень сильно контрастирует с громадным объемом имеющихся в настоящее [c.191]
Заметим, что различие по длине рестрикционных фрагментов между родителями и детьми может возникать не только в результате точковых мутаций в сайте узнавания, но и в результате ошибки репликации или кроссинговера. Ожидается, что эти события происходят чаще, чем точковые мутации. Среди 27 индивидов, у которых проанализировано 240 полос, выявлена такая полоса, которой не было ни у одного из родителей, отсюда скорость мутирования составляет 1/240, что по крайней мере на 4 порядка выше, чем скорость мутирования для точковых мутаций (разд. 2.3.3.9). [c.290]
Передача наследственной информации в неискаженном виде — важнейшее условие выживания как каждого конкретного организма, так и вида в целом. Следовательно, в ходе эволюции должна была сформироваться система, позволяющая клетке исправлять нарушения ДНК, вызванные ошибками репликации или повреждающими воздействиями окружающей среды. Подсчитано, что в результате повреждений, обусловленных этими причинами, в геноме клеток зародышевой линии человека происходит в среднем шесть нуклеотидных замен в год. По-видимому, в соматических клетках за год происходит примерно такое же число мутаций. [c.79]
В работе [59] было показано, что 95% мутаций, индуцируемых АП в Е.соЫ, относятся к транзициям А—Т — -> Г — Ц, т. е. преимущественно возникают ошибки репликации. [c.34]
Данные таблицы указывают, что в Si состоянии А и Т (У) легче, а Ц и Г труднее переходят в редкие формы, чем в состоянии Sq. Видно также, что в случае амино-иминной таутомерии А более склонен к таутомеризации, чем Ц лактам-лактимные превращения Т (У) более вероятны, чем Г. Таким образом, в состоянии Si положение противоположно тому, что имеется в So- Поэтому при действии УФ-света ошибки репликации чаще всего приводят к транзициям А—Т Г—Ц (рис. 3, а и б), а ошибки включения — к транзициям Г —Ц- А—Т (рис. 3, ж и з). Рассмотренный механизм объясняет возникновение мутаций, отнесенных к транзициям А — Т — Г — Ц [177]. [c.54]
ДНК-полимеразы проверяют комплементарность каждого нуклеотида матрице дважды один раз перед включением его в состав растущей цепи и второй раз перед тем, как включить следующий нуклеотид. Очередная фосфодиэфирная связь образуется лишь в том случае, если последний (З -концевой) нуклеотид затравки комплементарен матрице. Если же на предыдущей стадии полимеризации произошла ошибка (например, из-за того, что нуклеотид в момент полимеризации находился в необычной таутомерной форме), то репликация останавливается до тех пор, пока неправильный нуклеотид не будет удален. Некоторые ДНК-полимеразы обладают не только полимеризующей, но и 3 -экзонуклеазной активностью, «Которая отщепляет не спаренный с матрицей нуклеотид затравки. После чего полимеризация восстанавливается, от механизм, коррекция, заметно увеличивает точность работы ДНК-полимераз. Мутации, нарушающие З -экзонуклеазную активность ДНК-полимеразы, существенно повышают частоту возникновения прочих мутаций. Напротив, мутации, приводящие к усилению экзонуклеазной актив- ности относительно полимеризующей, снижают темп мутирования Генетического материала. [c.47]
Спонтанные генные мутации определяются ошибками при репликации ДНК, возникающими вследствие теплового движе-иия атомов и молекул. Очевидно, что ошибки транскрипции и трансляции не наследуются. [c.283]
Если ошибка синтеза не устраняется системами репарации, то неизбежна деформация дуплекса и искажение генетической программы. Такие сохраняющиеся при репликации изменения ДНК носят название мутации. Они могут быть спонтанными и индуцированными. Частота спонтанных мутаций невелика и составляет всего 10 —10 на клетку. В основном имеют место мутации, обусловленные действием внешних факторов физических (радиация), биологических (вирусы) и чужеродных химических веществ на генетический аппарат клеток. Наиболее многочисленными и опасными являются мутагены окружающей среды. Загрязнение воды и воздуха различными химическими отходами промышленных предприятий, химическими средствами защиты растений отрицательно сказывается на генетической программе всех живых организмов. В последние годы установлено, что ряд пищевых красителей, стабилизаторов и вкусовых добавок обладает выраженной мутагенной активностью, что привело к значительному ужесточению требований, связанных с применением химических веществ в пищевой промышленности. Многие лекарственные вещества также воздействуют на генетический аппарат клеток и должны подвергаться специальным генетическим испытаниям. [c.455]
Из уровня спонтанных мутаций у бактерий в расчете на одно поколение рассчитано, что вероятность одной репликационной ошибки при синтезе ДНК составляет порядка 10 . Эту величину можно рассматривать как отношение скоростей реакций правильной репликации [c.194]
Как обсуждалось в гл. 13, наследственная информация, заключенная в нуклеотидной последовательности ДНК, сохраняется неизменной благодаря действию сложных метаболических механизмов, обеспечивающих осуществление репликации и репарации. Мутации могут быть результатом ошибки на любом из многочисленных последовательных этапов этих процессов. Мутагенные факторы способны изменять как непосредственно структуру ДНК, так и структуру ферментов, прямо или косвенно участвующих в соответствующих метаболических процессах. Для понимания механизмов мутаций требуется знание нуклеотидной последовательности гена дикого типа и мутантного гена. Без этого невозможно понять связь между изменениями, происходящими в структуре ДНК и действием конкретных факторов или условий среды, вызывающих мутации. Современные методы клонирования генов сделали возможным прямое определение нуклеотидной последовательности ДНК. Однако еще совсем недавно при изучении молекулярной природы мутаций приходилось анализировать аминокислотные замены в белках, синтезируемых мутантными генами, а затем с помощью таблиц генетического кода выявлять изменения в нуклеотидной последовательности. [c.8]
Связь между ДНК, менделевскими факторами и признаками организма не столь проста, как представлялось прежде, но под наблюдения классических менделистов можно тем не менее подвести химическую основу. Установленные ранее явления доминантности и рецессивности рассматриваются теперь в таких аспектах, что можно без труда показать связь этих явлений с определенными химическими реакциями (разд. 2.1.2). Эпистаз (разд. 2.1.2) можно интерпретировать в понятиях взаимодействующих генов и генных продуктов. Нетрудно, например, представить себе, как может происходить химическое воздействие на экспрессию на уровне как самого гена, так и любого из ряда последовательных генных продуктов химическими факторами, содержащимися во внутренней и во внешней среде. Наконец, мутацию можно интерпретировать как ошибку в репликации ДНК. Такая ошибка может ограничиваться замещением одного нуклеотида другим (точковая мутация) или же выражаться в структурной перестройке целых хромосом (хромосомные мутации, в том числе нехватки, удвоения, транслокации и инверсии). [c.38]
Если УФ-свет вызывает мутацию гена, ответственного за биосинтез ДНК-полимеразы или других ферментов репарирующей системы, то возникшие мутантные клетки характеризуются высокой частотой спонтанных мутаций, которые не связаны с действием внешних или внутренних мутагенных факторов, а обусловлены ошибками в ходе репликации. В связи с этим в генетике возникло представление о гене, контролирующем частоту естественных мутаций,— гене-мута-торе. [c.311]
Устойчивость процесса отбора по отношению к систематическим помехам зависит прежде всего от уровня помех, т. е. от вероятности мутаций в единицу времени и вероятности ошибочной репликации. Следует учитывать также различие в характере повреждений, вызываемых помехами. При ошибочной редупликации (или в результате мутаций) могут возникнуть как летальные варианты, т. е. гиперциклы с весьма малой репродуктивной способностью, так и гиперциклы, способные к дальнейшему развитию, но отличающиеся от исходного. Характер повреждений зависит от стадии развития. На первой стадии (до возникновения адаптеров) все мутации и ошибки ведут к равноправным вариантам, летальных вариантов нет. На второй стадии, после образования адаптеров, но до возникновения единого кода летальные мутации существенной роли не играют, поскольку сводятся к уменьшению скорости репродукции. [c.36]
На наших глазах очередной кризис возник и в молекулярной генетике. Совсем недавно доминировали представления о необычайной точности и стабильности механизмов репликации и процессинга РНК, в результате чего мутации (ошибки репликации) возникают крайне редко, с частотой порядка 10 10 за одно поколение. Немало приводилось убедительных доводов в пользу того, что только такая надежность репликации ДНК и обеспечивает саму возможность прогрессивной эволюции — при учащении мутационного процесса отбор-де не справился бы с задачей поддержицання упорядоченности организации и совершевствовавяя [c.4]
По происхождению мутации делятся на спонтанные (неконтролируемые) и индуцированные (контролируемые). Первые возникают в результате неконтролируемого влияния каких-то естественных факторов (радиация, температура и т. д.). Направленное использование мутагенов приводит к возникновению индуцированных мутаций. Многими экспериментами четко показано, что мутации возникают независимо от условий среды обитания, т. е. не направленно. Мутации возникают в основном как ошибки репликации ДНК. Выделяют следующие типы мутаций перестройка хромосом, перестройка генома клетки грибов и водорослей (полиплоидия, гаплоидия, гетероплоидия), внутригенные изменения (прямые мутации, реверсии, обратные мутации). [c.102]
В качестве альтернативного подхода к измерению скоростей мутации А. Portner и соавт. [198] использовали моноклональные антитела и сравнили скорости мутации в отношении к резистентности антител к вирусам гриппа, везикулярного стоматита и Синдбис. Для всех трех вирусов скорости мутации составили около на репликацию, тогда как скорость ошибки репликации ДНК Е. соИ оценивалась как 10 —10 ° [63]. [c.190]
Спонтанные мутации возникают случайно, т.е. в любой. момент любой ген может претерпеть изменения. Причинами спонтанного мутационного процесса являются многочисленные факторы экзогенной и эндогенной природы, в том числе постоянное воздействие па организм человека мутагенов химической, биологической и физической природы (например, естественный фон облучения, действие вирусов) ошибки репликации ДНК, которые копируются и накапливаются в ряду клеточных поколений нарушение функционирования репаративных систем действие экзогенных метаболитов физи0 ю1ическ0е состояние и возраст организма. Спонтанные мутации могут возникать как в половых, так и в соматических клетках на генном, хромосомном и геномном уровнях. [c.125]
Разные аллели одного и того же Г. возникают благодаря мутациям-илслецуемьш изменениям в структуре исходного Г. В норме Г. чрезвычайно стабилен и при удвоении хромосом во время репликации ДНК воспроизводится совершенно точно вероятность ошибки не превышает 10″ . Мутации происходят редко и обычно влекут за собой неблагоприятные последствия для организма, т. к. нарушается его способность синтезировать нормальный белок. Однако в целом это явление играет положит, роль накопление редких полезных мутаций создает основу генетич. изменчивости, необходимой для эволюции. [c.517]
Равновесие между созидательными возможностями выбора среди специфических оснований в ДНК (созидательные мутации) и точностью синтеза белков (поддерживающих жизнь организма) является основой эволюции. Ферменты, которые заряжают тРНК специфической аминокислотой, обладают очень низкой вероятностью ошибки, порядка 1 Ю» для гомологичных аминокислот. При репликации точность даже выше, и величина ошибки редко превышает 1 на 10. [c.212]
Приближенная модель репликации ДНКизображена на рис. 2.11. Из приведенной схемы видно, что репликация точно воспроизводит прежнюю (исходную) структуру ДНК. Но если произошла ошибка в процессе копирования (мутация), то она будет с предельной точностью копироваться при последующих репликациях изменившейся ДНК. Показано, что участки ДНК, содержащие скопления нуклеотидов, обладают повышенной склонностью к спонтанным мутациям [22]. [c.94]
Для нормального функционирования аппарата исправления ошибок, связанных с включением неправильных нуклеотидов, необходимо располагать механизмом, позволяющим отличать новосинтезированную цепь ДНК от родительской матричной цепи. В противном случае с вероятностью 1/2 будет происходить исправление нуклеотида в родительской цепи, приводящее к закреплению потенциально мутагенной ошибки, допущенной ДНК-полимеразой. Вероятно, для установления различий между родительской и дочерней цепями ДНК в Е. соИ используется метилирование аденина в последовательности GAT . Эта палиндром-ная последовательность обычно метилирована в обеих цепях родительской ДНК. При полуконсервативной репликации метилированной ДНК образуется дочерняя ДНК, в которой одна цепь, пришедшая от родительской ДНК, метилирована, а новообразованная цепь в течение некоторого времени после выхода из области репликативной вилки остается неметилированной. Следует заметить, что метилирование новообразованной цепи ДНК осуществляется ферментом, отличным от метилаз, входящих в систему рестрикции—модификации, обсуждавшуюся в гл. 9. Бактерии dam , дефектные по метилированию аденина в результате нарушения синтеза соответствующей метилазы характеризуются повышенной частотой спонтанных мутаций, что подтверждает гипотезу об участии метилазы dam в системе исправления ошибок репликации. [c.123]
Анализ приведенных выше результатов дает возможность написать для преобладающих таутомерных форм оснований нуклеиновых кислот формулы, изображенные на фиг. 55. Минорные таутомерные формы, возможно, играют существенную роль в возникновении спонтанных мутаций, поскольку спаривание несоответствующих оснований (см. гл. ХУП1) должно привести к ошибке при включении оснований и при последующей репликации цепи. Можно показать, что если скорость включения основания в цепь нуклеиновой кислоты меньше скорости перехода минорного таутомера в доминирующую форму, то скорость спонтанных мутаций, обусловленных данным основанием, приблизительно равна константе равновесия между минорным и доминирующим таутомерами. К сожалению, для азо- [c.308]
Задание 189. Напишите программу для моделирования самоорганизации ДНК в качестве примера самоорганизуюшихся систем. Используйте для этого следующую простую модель. Пусть имеется 100 молекул ДНК, состоящих из 12 нуклеотидов четырех видов (их обозначим буквами А, Т, С и G). Последовательность нуклеотидов в этих 100 молекулах ДНК случайная. Назовем одну из последовательностей идеальной она имеет некоторые преимущества перед остальными. Из 100 молекул ДНК в результате репликации получается еще 100 молекул. Однако при репликации встречаются ошибки (мутации), например в количестве 1%. Теперь из 200 молекул 100 погибает. При этом имеет значение преимущество, которым обладают молекулы с последовательностью нуклеотидов, похожей на идеальную . (Например, при каждом совпадении нуклеотида и его положения в цепи с идеальной последовательностью вероятность гибели уменьшается в два раза.) Процессы репликации и гибели протекают очень быстро. В конце концов все молекулы ДНК должны получить идеальную последовательность нуклеотидов, хотя вероятность ее образования в результате случайного процесса составляет 1 16777216. Что будет, если мутации будут возникать чаще или реже [c.330]
Теперь мы уже вполне подготовлены к тому, чтобы приступить к вопросу, поставленному в гл. VU, а именно к вопросу о молекулярном механизме возникновения тех изменений в последовательности нуклеотидов ДНК, которые приводят к мутациям. Действительно, исследование характера возникновения мутаций Т-четных фагов с использованием методов генетического анализа с высоким разрешением дает большие возможности для проникновения в природу мутационного процесса. Использование фагов имеет еще одно важное преимущество по сравнению с ис-лользованием бактерий. Мутации фаговой ДНК можно изучать как в том случае, когда она находится в состоянии покоя вне клетки в составе инфекционной фаговой частицы, так и когда она находится в реплицирующемся, внутриклеточном, вегетативном состоянии. Уже самые первые исследования Херши и Лурия показали, что частота спонтанных мутаций в покоящейся ДНК очень мала — столь мала, что в течение многих лет считалось (как потом оказалось, ошибочно), что внеклеточные фаговые частицы вообще не мутируют месяцами и даже годами. Таким образом, новые мутации появляются в основном во время вегетативного размножения фага в клетке-хозяине. Рассмотрим следующий пример. Культуру Е. oli заражают препаратом фага Т2/- с титром 10 частица/мл. Фагу дают размножиться в течение нескольких циклов, пока все бактерии в культуре не подвергнутся лизису, а титр фага не достигнет величины 10 частица/мл. Оказывается при этом, что с каждым циклом размножения доля г-мутантов во всей популяции фагов увеличивается (примерно с 10″ в начале до 10 в конце). Следовательно, мутанты фага возникают в результате ошибок копирования при внутриклеточной репликации его генетического материала. Репликация ДНК родительского фага является очень точным процессом. И все же при репликации иногда происходит ошибка, порождающая в одной из вегетативных реплик изменение последовательности нуклеотидов, или мутацию. Мутантная реплика генетического материала включается затем при созревании в инфекционную фаговую частицу, которая в свою очередь заражает новую бактериальную клетку. В этой клетке очень точно копируется уже измененная информация, содержащаяся в мутантной частице поэтому все потомство такой частицы оказывается тоже мутантным. Поскольку репликация ДНК вегетативного фага происходит в соответствии с постулированным Уотсоном и Криком полуконсервативным механизмом, размножение фагового генома можно рассматривать как процесс бинарного деления и с точки зрения статистического анализа совершенно аналогичным процессу размножения генома бактерий. Следовательно, уравнение, связывающее долю мутантных особей п среди общего числа N потомков одного исходного родителя, возникших после g генераций, с частотой мутаций а [c.315]
Если элементарное мутационное событие представляет собой [включение неправильного нуклеотида в определенный участок синтезируе-мой полинуклеотидной реплики и если ДНК вегетативного фага реплицируется в соответствии с полуконсервативным механизмом Уотсона и Крика, то мы можем предсказать такую особенность вновь рождаюш егося мутантного генома, которую без знания молекулярной основы процесса мутирования вообще невозможно было бы предвидеть. Предположим, что во время синтеза цепи-реплики происходит одна из редких ошибок копирования, например остаток тимина в родительской цепи незаконно спаривается с гуанином, а не с аденином. В результате этого мутагенного акта репликации возникает двойная спираль с исходной ин-формацией в старой (родительской) цепи и мутантной информацией в цепи, синтезированной заново (фиг. 160). При следующем цикле репликации комплементарные нити этой мутантной молекулы вновь разъединяются и каждая из них, функционируя как матрица, синтезирует новую комплементарную цепь. В результате появляется одна двойная спираль ДНК, несущая мутантную информацию в обеих цепях, и одна немутантная двойная спираль. Исходная мутантная молекула ДНК представляет собой, следовательно, гетеродуплексную гетерозиготу, которая несет в одном участке два аллеля — мутантный и немутантный, по которым при следующем цикле репликации происходит расщепление. Можно ожидать, что во время внутриклеточного размножения фага некоторые молекулы ДНК фага с мутацией, возникшей в результате ошибки копирования при последней репликации, будут извлечены из вегетативного фонда фага и войдут в состав зрелых инфекционных частиц. Эти частицы и будут мутационными гетерозиготами. [c.325]
Во-первых, исходным повреждением, отвечающим за потенциальный мутагенный эффект, должны быть тиминовые димеры. Во-вторых, процессом, превращающим потенциальные изменения в окончательные мутации, не может быть репарация за счет иссечения и заполнения (она могла бы приводить к мутации, если бы, например, точность репарационной репликации, изображенной на фиг. 187, была невелика и допускала ошибки копирования). Можно заключить, следовательно, что мутация вызывается нерепарированными тиминовыми димерами в тех клетках, которые выжили, несмотря на наличие такого неисправленного п овреж- [c.382]
Другой путь возникновения транзиций-это случаи ошибочного спаривания, приводящие к возникновению неканонических пар и, следовательно, к дефектам в уотсон-криковской спирали. В нормальном цикле репликации такая ошибка может случайно произойти вследствие включения неправильного основания. Спонтанная частота ошибок определяется прежде всего точностью фермента ДНК-полимеразы, отвечающей за репликацию (см. гл. 32). Существует также более ограниченный репара-тивный синтез ДНК, который активируется в результате генетической рекомбинации или повреждения ДНК (см. гл. 34). Различные системы репарации характеризуются разной частотой ошибок. Например, одна из репара-тивных систем Е. соИ особенно часто делает ошибки, и, следовательно, ее активация может стимулировать образование мутаций. Мы не располагаем достаточной информацией о частоте возникновения мутаций такого рода. [c.38]
Что представляет собой механизм появления ошибок Можно предположить, что определенный компонент пути репарации обусловливает продолжение репликации за сайтом повреждения. Когда ДНК-полимераза минует тиминовый димер, она включает неправильные основания и это приводит к появлению мутации. Существуют доказательства того, что для индукции ошибок необходимо присутствие ДНК-полимеразы III, обычной репликазы. Следовательно, рассматриваемая функция действует согласованно с нормальным реплика-ционным аппаратом. Мутации в гене, получившем название итиС, устраняют УФ-индуцируемый мутагенез, но не нарушают какие-либо известные ферментативные функции. Вероятно, продукт этого гена, итиС, служит компонентом системы, продуцирующей ошибки. [c.440]
Модель Уотсона-Крика позволяет представить себе, как может удваиваться нативная молекула ДНК, образуя две одинаковые дочерние молекулы. Поскольку две цепи ДНК комплементарны, каждая из них при расплетании двойной спирали может служить матрицей для синтеза новой комплементарной цепи. Последовательность оснований во вновь синтезируемой цепи будет определяться спецификой водородных связей между основаниями цепи-щаблона и вновь образуемой цепи (рис. 4.13). Таким образом, генетическая информация, содержавшаяся в последовательности пар оснований родительской молекулы, будет полностью воспроизведена в двух дочерних молекулах. Более того, если в процессе удвоения ДНК произошла ошибка и какой-то нуклеотид во вновь образуемой цепи выпал или оказался некомплементарным исходному, то это может изменить информационное содержание молекулы, причем можно ожидать, что эта ошибка будет передана дочерним молекулам ДНК в следующих поколениях. Такая замена пары нуклеотидов может обладать свойствами генетических мутаций. Таким образом, модель структуры ДНК Уотсона и Крика объясняет как способность генов к самоудвоению (репликации), так и их информационные свойства. [c.107]
Несмотря на корректорские функции, присущие ДНК-полимеразам Е. соИ, некоторые нуклеотиды оказываются все же ошибочно включенными в новообразованную цепь ДНК. Их присутствие делает возможным возникновение спонтанных мутаций, в том случае если ошибки не будут исправлены до начала следующего цикла репликации. Свидетельства в пользу существования пострепликационных систем исправления ошибок, или репарации, были получены при изучении таких явлений, как [c.122]
Спонтанные трацзиции могут происходить при репликации ДНК вследствие таутомеризации, т. е. изменения положения протона, меняющего химические свойства молекулы. Таутомеризация в нуклеотидных основаниях меняет их способность образовывать водородные связи, так что аденин приобретает свойства гуанина, гуанин-аденина, цитозин-тимина, а тимин-цитозина (рис. 20.2). Мутагенная активность 5-бром-урацила, аналога тимина, в котором метиловая группа замещена атомом брома, обусловлена таутомеризацией, связанной с большим, нежели у метиловой группы, сродством к электрону атома брома по сравнению с метиловой группой (рис. 20.3). Индуцируемые 5-бромура-цилом мутации могут обусловливаться либо ошибками при включении, [c.9]
Gene mutations can’t really turn baby turtles into cartoon super heroes like the Teenage Mutant Ninja Turtles.
Genetic mutations are slight alterations of DNA or RNA nucleotides, genes or chromosomes that may occur during replication or cell division. Random, uncorrected errors may be beneficial or harmful in relationship to evolution.
Some effects of gene mutation go unnoticed.
What Is Gene Mutation in Biology?
The two types of mutations in biology mainly occur in germ (egg and sperm) cells and in somatic (body) cells.
Germline mutations that give rise to genetic disorders can be inherited due to alterations in DNA sequences. Somatic mutations like lung cancer associated with heavy smoking can’t be passed down to subsequent generations.
Different mutations can prove deadly to an organism if gene regulation is severely disrupted. On the other hand, random mutations may give organisms with that mutated trait a competitive advantage.
For example, Charles Darwin found a correlation between the beak shape of finches and their prevalence in divergent habitats on the Galapagos Islands. Darwin’s work led to the theory of natural selection.
When Do Gene Mutations Occur?
Mutations frequently occur just before the process of mitosis when DNA is being replicated in the cell nucleus. During mitosis or meiosis, mishaps can occur when chromosomes are not lined up correctly or fail to separate properly. Chromosomal mutations in the germ cells can be inherited and passed along to the next generation.
Some gene mutations can interfere with the rate of normal cell growth and increase cancer risk. Mutations in non-reproductive cells can trigger benign growths or cancerous tumors such as melanoma in skin cells. Defective genes on chromosomes are passed on, as well as too many or too few chromosomes per cell when these mutations happen in germline cells.
Gene mutation examples include severe genetic disorders, cell overgrowth, tumor formation and heightened risk of breast cancer. Cells have a finely tuned mechanism for correcting mutations at checkpoints during cell division, which detects most mutations. Once DNA proofreading is completed, the cell proceeds to the next stage of the cell cycle.
Causes of Gene Mutations
Mutations can occur because of external factors, also known as induced mutations. Mutagens are external factors that can cause alterations to DNA. Examples of potentially harmful environmental factors include toxic chemicals, X-rays and pollution. Carcinogens are mutagens that cause cancer such as UV radiation.
Various types of spontaneous mutations happen due to mistakes in cell division or reproduction, as well as during DNA replication or transcription. During DNA replication, nucleotide bases can be added or deleted, or a segment of DNA may be translocated to the wrong place on a chromosome.
When the cell is dividing, mistakes can occur during chromosomal separation, resulting in abnormal numbers and types of chromosomes with different genes. Such mutations can also be passed on from parent to child.
Types of Gene Mutations
The genetic code determines the order of codons that will create building blocks of amino acids and proteins. Mutations frequently occur, which isn’t surprising given the billions of cells in the body that are perpetually dividing to replace old, worn out cells.
Most of the time, errors in DNA replication or segregation are quickly repaired by enzymes or the cell is destroyed before they can cause lasting damage. When DNA repair attempts fail, spontaneous mutations stay within the DNA. Benign spontaneous mutations increase the genetic variance and biodiversity of a population.
The following are some of the types of gene mutations that can occur:
- Tautomerism: This occurs during replication of DNA in the cell nucleus. Tautomers are mismatched pairs of nucleotide bases.
- Depurination: This is a chemical reaction that happens when the bonds break between the deoxyribose sugar in DNA and the purine base of guanine or adenine. Losing a purine base is a common spontaneous mutation.
- Deamination: This occurs if enzymes remove a nitrogen group from an amino acid. For instance, the (temperature-dependent) hydrolic deamination of cytosine to uracil is a leading cause of single-site, spontaneous mutations, as reported in the Proceedings of the National Academy of Sciences.
- Transition and transversion: These mutations are two types of DNA substitution errors that involve switching of nucleotide base pairs.
- Transition: This occurs due to a genetic shuffling of similarly shaped nucleotide bases. A transition mutation occurs when a wild-type (normally occurring) base pair like adenine and thymine are replaced by guanine and cytosine base pairs.
- Transversion: This refers to the interchange of differently shaped purine and pyrimidine bases. For example, the mutated segment of DNA may have adenine replacing thymine.
Types of Point Mutations
Changes in the number or type of nucleotides are called point mutations. The effects of point mutation can range from harmless to life threatening. Mispairing or reordering of nucleotide bases are considered silent mutations when the change doesn’t affect cell functioning. The new amino acid may even perform the same functions as the one it replaced.
The following are types of point mutations that can occur:
- Missense mutation: This happens when one nucleotide is replaced with another. Substitutions of bases can interfere with normal protein syntheses and functioning. For instance, a single point mutation on the hemoglobin beta (HBB) gene causes sickle cell anemia blood disorders.
- Nonsense mutations: These occur when atypical base pairings produce a stop codon that may cause improper functioning or impede functioning altogether.
- Frameshift mutations: These are point mutations that result when a nucleotide pair is added or omitted in a gene sequence that shifts how codons are read. Such mutations often result in different amino acids being added to the protein being synthesized. An example is beta thalassemia, a blood disorder caused by mutations to the HBB gene. Diseases like cystic fibrosis involve gene mutation deletion – when nucleotides, amino acids, base pairs or whole genes are removed.
Copy Number Variation Mutations
Gene amplification is involved in the production of extra copies of genes with heightened expression. Duplication or amplification is seen in some breast cancers and other types of malignancies, for instance. Overproduction of repeated codons in a gene alters gene functioning.
For instance, Fragile X syndrome is an intellectual disability caused by a high number of trinucleotide repeats that impair DNA stability.
Gene Mutations and Chromosomal Mutations
Significant mutations can result when the structure or number of chromosomes change. Chromosomal aberrations may occur during mitosis or meiosis.
Mutations can also involve sex chromosomes X and Y and can affect gender expression.
Gene Mutation Diseases
Errors in meiosis can result in deletion of chromosomal segments. For instance, cri du chat syndrome results from a missing piece of genetic material on the arm of chromosome 5. When a part of a chromosome breaks off, it may attach to another chromosome.
The following are a few examples:
- Duplications or amplifications: These happen when a chromosome is added to a homologous chromosome that already contains that sequence, as seen in some cancers.
- Inversions: These occur when part of chromosome breaks off and then reattaches backwards. For instance, Optiz-Kaveggia syndrome is linked to this type of mutation.
- Translocation: This is when a component of a chromosome attaches to a non-homologous chromosome. A form of leukemia is associated with translocation mutation.
- Nondisjunction: This is the failure of chromosomal separation, causing reproductive cells to have too many or two few chromosomes. Possible consequences can include miscarriage, Down syndrome and Turner syndrome.
Mutations and Genetic Counseling
Prenatal diagnosis for high-risk populations and other types of genetic counseling, including DNA genetic testing kits, can provide helpful medical information for family planning. Screening and early detection leads to better treatment outcomes. There are also genetic benefits for certain disease carriers that are good to know.
Carriers of the sickle cell gene have a protective factor against malaria, which is particularly advantageous in tropical regions. Research suggests a similar evolutionary advantage to being a carrier of cystic fibrosis and cholera resistance. Carriers of the cystic fibrosis gene may be better able to retain fluid and recover if exposed to cholera, according to preliminary studies.
Gene mutations can’t really turn baby turtles into cartoon super heroes like the Teenage Mutant Ninja Turtles.
Genetic mutations are slight alterations of DNA or RNA nucleotides, genes or chromosomes that may occur during replication or cell division. Random, uncorrected errors may be beneficial or harmful in relationship to evolution.
Some effects of gene mutation go unnoticed.
What Is Gene Mutation in Biology?
The two types of mutations in biology mainly occur in germ (egg and sperm) cells and in somatic (body) cells.
Germline mutations that give rise to genetic disorders can be inherited due to alterations in DNA sequences. Somatic mutations like lung cancer associated with heavy smoking can’t be passed down to subsequent generations.
Different mutations can prove deadly to an organism if gene regulation is severely disrupted. On the other hand, random mutations may give organisms with that mutated trait a competitive advantage.
For example, Charles Darwin found a correlation between the beak shape of finches and their prevalence in divergent habitats on the Galapagos Islands. Darwin’s work led to the theory of natural selection.
When Do Gene Mutations Occur?
Mutations frequently occur just before the process of mitosis when DNA is being replicated in the cell nucleus. During mitosis or meiosis, mishaps can occur when chromosomes are not lined up correctly or fail to separate properly. Chromosomal mutations in the germ cells can be inherited and passed along to the next generation.
Some gene mutations can interfere with the rate of normal cell growth and increase cancer risk. Mutations in non-reproductive cells can trigger benign growths or cancerous tumors such as melanoma in skin cells. Defective genes on chromosomes are passed on, as well as too many or too few chromosomes per cell when these mutations happen in germline cells.
Gene mutation examples include severe genetic disorders, cell overgrowth, tumor formation and heightened risk of breast cancer. Cells have a finely tuned mechanism for correcting mutations at checkpoints during cell division, which detects most mutations. Once DNA proofreading is completed, the cell proceeds to the next stage of the cell cycle.
Causes of Gene Mutations
Mutations can occur because of external factors, also known as induced mutations. Mutagens are external factors that can cause alterations to DNA. Examples of potentially harmful environmental factors include toxic chemicals, X-rays and pollution. Carcinogens are mutagens that cause cancer such as UV radiation.
Various types of spontaneous mutations happen due to mistakes in cell division or reproduction, as well as during DNA replication or transcription. During DNA replication, nucleotide bases can be added or deleted, or a segment of DNA may be translocated to the wrong place on a chromosome.
When the cell is dividing, mistakes can occur during chromosomal separation, resulting in abnormal numbers and types of chromosomes with different genes. Such mutations can also be passed on from parent to child.
Types of Gene Mutations
The genetic code determines the order of codons that will create building blocks of amino acids and proteins. Mutations frequently occur, which isn’t surprising given the billions of cells in the body that are perpetually dividing to replace old, worn out cells.
Most of the time, errors in DNA replication or segregation are quickly repaired by enzymes or the cell is destroyed before they can cause lasting damage. When DNA repair attempts fail, spontaneous mutations stay within the DNA. Benign spontaneous mutations increase the genetic variance and biodiversity of a population.
The following are some of the types of gene mutations that can occur:
- Tautomerism: This occurs during replication of DNA in the cell nucleus. Tautomers are mismatched pairs of nucleotide bases.
- Depurination: This is a chemical reaction that happens when the bonds break between the deoxyribose sugar in DNA and the purine base of guanine or adenine. Losing a purine base is a common spontaneous mutation.
- Deamination: This occurs if enzymes remove a nitrogen group from an amino acid. For instance, the (temperature-dependent) hydrolic deamination of cytosine to uracil is a leading cause of single-site, spontaneous mutations, as reported in the Proceedings of the National Academy of Sciences.
- Transition and transversion: These mutations are two types of DNA substitution errors that involve switching of nucleotide base pairs.
- Transition: This occurs due to a genetic shuffling of similarly shaped nucleotide bases. A transition mutation occurs when a wild-type (normally occurring) base pair like adenine and thymine are replaced by guanine and cytosine base pairs.
- Transversion: This refers to the interchange of differently shaped purine and pyrimidine bases. For example, the mutated segment of DNA may have adenine replacing thymine.
Types of Point Mutations
Changes in the number or type of nucleotides are called point mutations. The effects of point mutation can range from harmless to life threatening. Mispairing or reordering of nucleotide bases are considered silent mutations when the change doesn’t affect cell functioning. The new amino acid may even perform the same functions as the one it replaced.
The following are types of point mutations that can occur:
- Missense mutation: This happens when one nucleotide is replaced with another. Substitutions of bases can interfere with normal protein syntheses and functioning. For instance, a single point mutation on the hemoglobin beta (HBB) gene causes sickle cell anemia blood disorders.
- Nonsense mutations: These occur when atypical base pairings produce a stop codon that may cause improper functioning or impede functioning altogether.
- Frameshift mutations: These are point mutations that result when a nucleotide pair is added or omitted in a gene sequence that shifts how codons are read. Such mutations often result in different amino acids being added to the protein being synthesized. An example is beta thalassemia, a blood disorder caused by mutations to the HBB gene. Diseases like cystic fibrosis involve gene mutation deletion – when nucleotides, amino acids, base pairs or whole genes are removed.
Copy Number Variation Mutations
Gene amplification is involved in the production of extra copies of genes with heightened expression. Duplication or amplification is seen in some breast cancers and other types of malignancies, for instance. Overproduction of repeated codons in a gene alters gene functioning.
For instance, Fragile X syndrome is an intellectual disability caused by a high number of trinucleotide repeats that impair DNA stability.
Gene Mutations and Chromosomal Mutations
Significant mutations can result when the structure or number of chromosomes change. Chromosomal aberrations may occur during mitosis or meiosis.
Mutations can also involve sex chromosomes X and Y and can affect gender expression.
Gene Mutation Diseases
Errors in meiosis can result in deletion of chromosomal segments. For instance, cri du chat syndrome results from a missing piece of genetic material on the arm of chromosome 5. When a part of a chromosome breaks off, it may attach to another chromosome.
The following are a few examples:
- Duplications or amplifications: These happen when a chromosome is added to a homologous chromosome that already contains that sequence, as seen in some cancers.
- Inversions: These occur when part of chromosome breaks off and then reattaches backwards. For instance, Optiz-Kaveggia syndrome is linked to this type of mutation.
- Translocation: This is when a component of a chromosome attaches to a non-homologous chromosome. A form of leukemia is associated with translocation mutation.
- Nondisjunction: This is the failure of chromosomal separation, causing reproductive cells to have too many or two few chromosomes. Possible consequences can include miscarriage, Down syndrome and Turner syndrome.
Mutations and Genetic Counseling
Prenatal diagnosis for high-risk populations and other types of genetic counseling, including DNA genetic testing kits, can provide helpful medical information for family planning. Screening and early detection leads to better treatment outcomes. There are also genetic benefits for certain disease carriers that are good to know.
Carriers of the sickle cell gene have a protective factor against malaria, which is particularly advantageous in tropical regions. Research suggests a similar evolutionary advantage to being a carrier of cystic fibrosis and cholera resistance. Carriers of the cystic fibrosis gene may be better able to retain fluid and recover if exposed to cholera, according to preliminary studies.